DEMO* Lomvi
Engelsk navn: Common guillemot
Vitenskapelig navn: Uria aalge
Beskrivelse
Lomvi er den største av alkefuglene. Den kjennetegnes ved at den hekker i tette kolonier i fuglefjellene, hvor den legger sitt ene egg rett på åpne hyller eller i steinurer.
Fuglen er ca 45 cm lang og veier rundt 1 kg. Fjærdrakten er brunsvart og hvit, hvor hode, nakke og strupe, rygg og overside av vinger er brunsvarte, og bryst og buk er hvit. Til forskjell fra sin slektning polarlomvien som den ligner mye på, har lomvien mørke, melerte tegninger på kroppsidene.
Et annet kjennetegn som skiller lomvien fra polarlomvi er at den er større med et spinklere og litt lengre nebb. Hunnen og hannen er like i utseende, men hannen er en anelse større og tyngre enn hunnen.
Lomvien finnes i to ulike fargevarianter (morfer), hvor den ene kjennetegnes med en hvit ring rundt øyet som fortsetter som en hvit stripe bakover mot øret. Det ser ut som den har briller eller en ring rundt øyet, og den kalles ringvi på norsk. Den andre fargevarianten mangler denne hvite «brillen» og er helt mørk på hele hodet, inkludert rundt øyet.
Sommerdrakten til lomvien kjennetegnes ved at hele hodet og halsen er mørk. I vinterdrakt er kinn, strupe og hals hvite.
Taxonomisk befinner lomvien seg i ordenen Charadriiformes (Vade-, måke- og alkefugler) og familien Alcidae (Alkefamilien). Det finnes flere underarter, og i nordlige områder, inkludert våre strøk, finnes U. a. aalge, mens U. a. intermedia er mer vanlig i Østersjøen (Pontoppidan, 1763).
Relevante lenker:

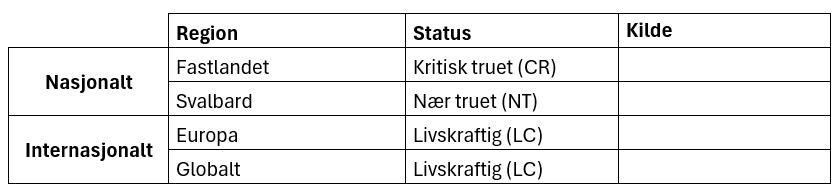
Rødlistestatus for lomvi
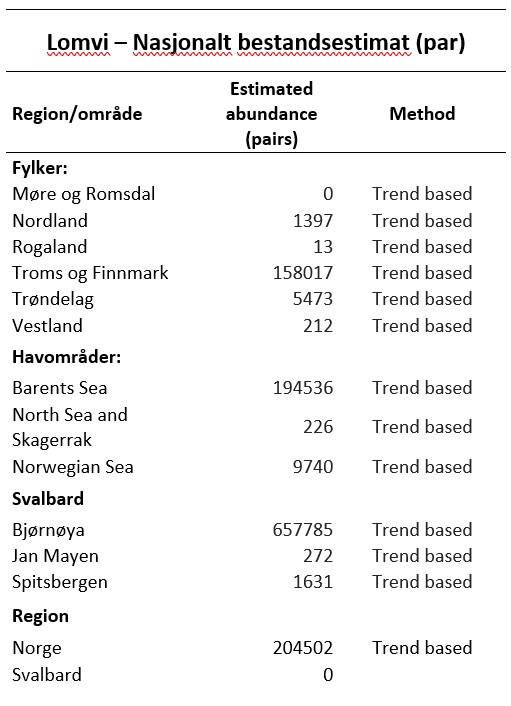
Bestandsstørrelse
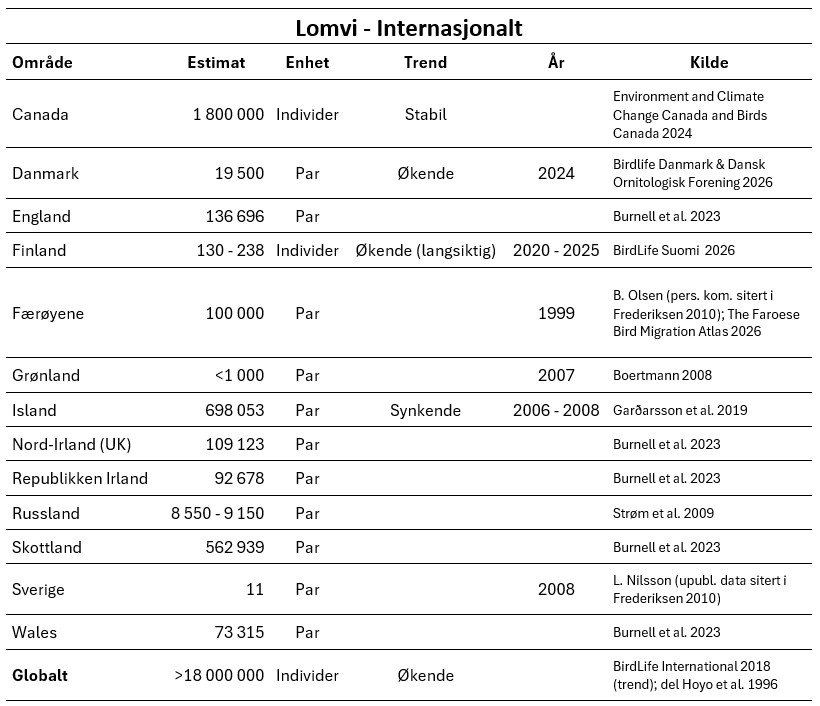
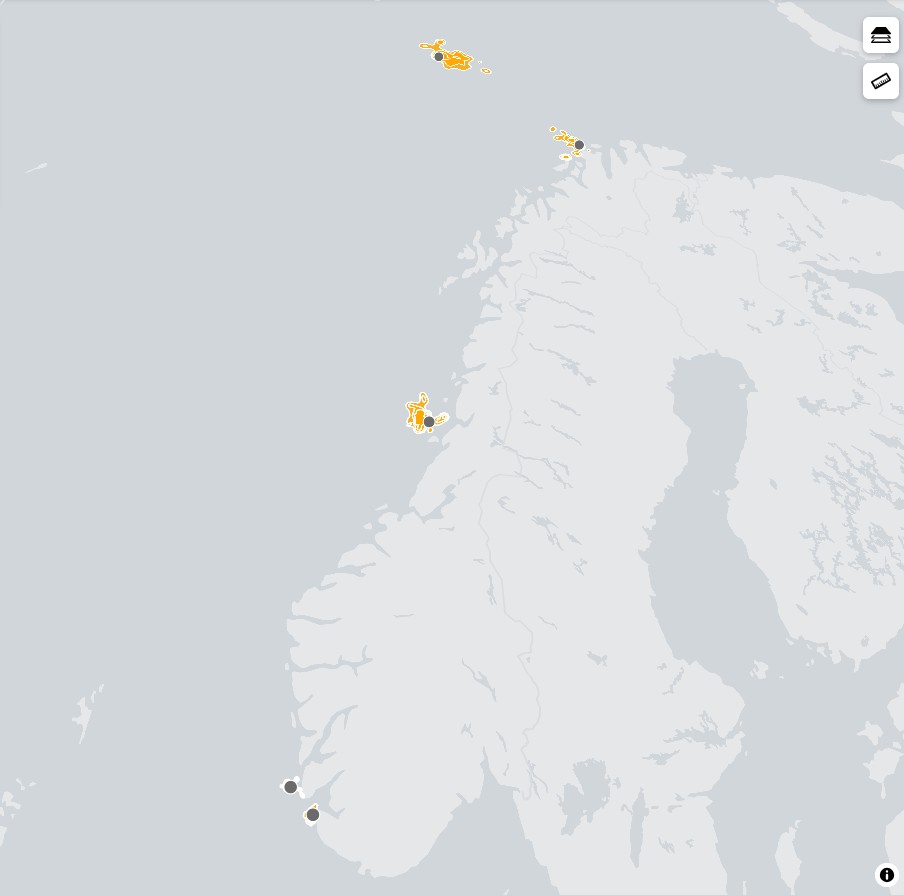
Hekkeutbredelse
Lomvien hekker rundt hele den nordlige halvkule (Gaston og Jones 1998), og det finnes hekkekolonier både i Nord-Atlanteren og i det nordlige Stillehavet. Sørligste hekkeforekomst i Europa er Portugal, og nordligste er Svalbard og Novaja Semlja (Russland). Det finnes også hekkeforekomster i Baltikum.
Lomvien har en mer sørlig, såkalt boreal utbredelse enn den mer arktiske polarlomvien, men i enkelte kolonier finnes det overlapp mellom begge artene. I Norge finnes hekkeforekomster fra Rogaland i sør til Øst-Finnmark i nord, i tillegg til hekkeforekomstene på Jan Mayen, Bjørnøya og Spitsbergen.
Trekk og vinterområder
I Norge har vi god kunnskap om trekk og vinter-utbredelsen til lomvi fra nøkkellokaliteter hvor hekkende individer har vært utstyrt med helårlige sporingsbrikker (Sklinna, Hjelmsøya, Hornøya og Bjørnøya) (SEATRACK.net).
Om høsten trekker nesten alle lomviene fra disse koloniene til Barentshavet. Høstutbredelsen til disse koloniene overlapper en god del, og det sentrale eller sør-østlige Barentshavet utpeker seg som et viktig høst-område da disse bestandene utgjør ca 90% av norges totale hekkebestander av lomvi.
Rett etter hekketiden myter lomvien. Siden den blir flyveudyktig i deler av mytetida har den behov for å være i et område med forutsigbare og gode næringsforhold. Barentshavet er kjent for å være svært næringsrikt om høsten (Wassmann et al. 2006).
I tillegg har lomvien et svømmetrekk, hvor hannen svømmer sammen med ungen fra hekkeplassen til et oppvekstområde. Ungen forlater hekkeplassen når den er 1/3 av voksen størrelse og fortsatt flyveudyktig. Det er vist at svømmetrekket fra Bjørnøya og Hornøya går fra kolonien til et område i det sørøstlige Barentshavet (Erikstad et al upublisert, Merkel og Strøm 2024), og det ser ut som de utnytter havstrømmer for å effektivisere svømmetrekket.
Vinterutbredelsen til ulike lomvibestander varierer litt mer (SEATRACK.net). Lomvi fra Bjørnøya, Hornøya og Hjelmsøya forblir i Barentshavet gjennom vinteren, men har mindre grad av overlapp med hverandres utbredelse.
Hornøya-fuglene holder seg i det sørøstlige delene av Barentshavet, mens fuglene fra Hjelmsøya trekker litt mer vestover og de som hekker på Bjørnøya holder seg mer i det sentrale Barentshavet. Lomvi fra Sklinna trekker i hovedsak ut av Barentshavet og sørover tilbake mot kysten av Midt-Norge.
Habitatbruk i hekkesesongen
Tekst kommer. Kartet skal kunne zoomes og panoreres
Bestandsutvikling
Ifølge Brun (1969) var den norske lomvibestanden på 1960-tallet estimert til 240 000 – 320 000 individer.
I tiårene etterpå gikk lomvibestandene sterkt tilbake, og perioden 1980 til 2013 viste en bestandsnedgang på ca. 80 – 90 % (Fauchald et al. 1990). I 1986/87 skjedde en stor akutt bestandsreduksjon i bestandene rundt Barentshavet, noe som i ettertid har vist seg å skyldes sammenfallende lave forekomster av flere viktige byttedyrbestander, som lodde og yngre årsklasser av sild og torsk (Erikstad et al. 2013), og spesielle vær- og klimaforhold (Mesquita et al. 2015).
Ifølge estimeringer for perioden 2013 til 2024, er det variasjon i utviklingen av bestandsstørrelse mellom fastlandskoloniene, mens det er en økning i hekkebestandene på Bjørnøya og Spitsbergen.
Sklinna og Røst har hatt økning fra hhv ca 1000 til 3500 individer og 43 til 99 individer det siste tiåret. Mens Runde, Hjelmsøya og Hornøya har hatt nedgang, med hhv 101 til 7 individer (Runde), 13 500 til 4700 individer (Hjelmsøya) og 24 000 til 12 000 individer (Hornøya). På Bjørnøya har antallet hekkende lomvier økt fra ca 203 000 til ca 290 000 individer.
Nøkkellokaliteter
Demografi
Overlevelse
Som alle sjøfugler lever lomvien lenge. Den lengstlevende lomvi registrert er en lomvi i Sverige på 49 år. Gjennomsnittlig levealder er 23 år. Den høyeste alder som er registrert i Norge er en lomvi på Hornøya som ble ringmerket som voksen hekkefugl i 1988, og som ble sett igjen hekkende på samme fjellhylle sommeren 2025 (Reiertsen upublisert). Dersom en antar at første år som hekkefugl er 4 år, vil denne være minimum 42 år.
Reproduksjon
Siden lomvi lever lenge, er også voksenoverlevelsen høy for denne arten (gjennomsnitt i Norge = 91,3%).
Lomvien legger kun ett egg og fostrer derfor opp maksimalt en unge per år. Reproduksjonen regnes derfor som lav, og overlevelsen til unger varierer mye fra år til år. Gjennomsnittlig alder for å starte reproduksjon er 4 år, men kan variere fra 2 – 7 år (Ainley et al. 2021).
Timing for egglegging og klekking kan være en driver for demografien til arter, og kan være påvirket av klimaendringer. Men dykkende fugler som lomvi, som også har en relativt kort hekkesesong, har vist mindre grad av endring i timing enn andre sjøfuglarter (Descamps et al. 2019). Eksempelvis har ankomstdatoen for lomvi på Hornøya vist seg å være i perioden fra tidlig mars til medio mars (Reiertsen upublisert), egglegging starter jevnt over fra rundt 20. mai (Reiertsen pers obs) og klekking starter rundt 20. juni, med liten variasjon i gjennomsnittlig klekkedato mellom år (Burr et al. 2016).
Næringsvalg
Lomvien regnes som en fiskespiser, og spiser stimfisk som lodde, sild, og tobis, men også yngre årsklasser av for eksempel sei og torskefisk (Barrett et al 1997). Den dykker etter maten sin, og bruker vingene for å manøvrere under vann. Vinterstid kan dykkene gå ned til mer enn 150 meter.
I hekketiden bringer den en og en fisk inn til ungen sin, som den holder i nebbet. Det er vist at voksne hekkefugler foretrekker å spise torskelarver (Bugge et al. 2010), som er vist å drifte med kyststrømmer fra gytegrunner lenger sør langs kysten, og akkumulere ved fuglefjellene i nord (Sandvik et al. 2016).
Påvirkningsfaktorer og trusler
Ifølge Fauchald et al. 2015 og Birdlife International (2020) er truslene mot lomvi-bestandene flere, og inkluderer endring in næringstilgang som følge av klimaendringer og overfiske, bifangst fra fiskerier, oljesøl og forurensing, predasjon og forstyrrelser i hekketiden.
Referanser
Ainley, D. G., D. N. Nettleship, and A. E. Storey (2021). Common Murre (Uria aalge), version 2.0. In Birds of the World (S. M. Billerman, P. G. Rodewald, and B. K. Keeney, Editors). Cornell Lab of Ornithology, Ithaca, NY, USA. https://doi.org/10.2173/bow.commur.02
Anker-Nilssen, T., Barrett, R.T., Lorentsen, S.-H., Strøm, H., Bustnes, J.O., Christensen-Dalsgaard, S., Descamps, S., Erikstad, K.E., Fauchald, P., Hanssen, S.A., Lorentzen, E., Moe, B., Reiertsen, T.K. & Systad, G.H. (2015). SEAPOP. De ti første årene. Nøkkeldokument 2005-2014. SEAPOP. Norsk institutt for naturforskning, Norsk Polarinstitutt & Tromsø Museum – Universitetsmuseet. Trondheim, Tromsø. 58 s.
Artsdatabanken: Lomvi Uria aalge (Pontoppidan, 1763) Hentet 09.03.2026 fra https://artsdatabanken.no/arter/takson/3570
Barrett, R. T., Bakken, V., & Krasnov, J. V. (1997). The diets of common and Brünnich’s guillemots Uria aalge and U. lomvia in the Barents Sea region. Polar research, 16(2), 73-84.
Bird, J. P., Martin, R., Akçakaya, H.R., Gilroy, J., Burfield, I.J., Garnett, S., Symes, A., Taylor, J., Sekercioglu, Ç.H. & Butchart, S.H. (2020). Generation lengths of the world’s birds and their implications for extinction risk. Conservation Biology
BirdLife International (2020) IUCN Red List for birds. Downloaded from http://www.birdlife.org on 26/08/2020
Bugge, J., Barrett, R.T. & Pedersen, T. Optimal foraging in chick-raising Common Guillemots (Uria aalge). J Ornithol 152, 253–259 (2011). https://doi.org/10.1007/s10336-010-0578-9
Burr, Z. M., ØVarpe, T.Anker-Nilssen, K. E.Erikstad, S.Descamps, R. T.Barrett, C.Bech, S.Christensen-Dalsgaard, S.-H.Lorentsen, B.Moe, T. K.Reiertsen, and H.Strøm. 2016. Later at higher latitudes: large-scale variability in seabird breeding timing and synchronicity. Ecosphere7(5):e01283. 10.1002/ecs2.1283
Descamps S, Ramírez F, Benjaminsen S, et al. Diverging phenological responses of Arctic seabirds to an earlier spring. Glob Change Biol. 2019;25:4081–4091. https://doi.org/10.1111/gcb.14780
Erikstad KE, Reiertsen TK, Barrett RT, Vikebø F, Sandvik H (2013). Seabird–fish interactions: the fall and rise of a common guillemot Uria aalge population. Mar Ecol Prog Ser 475:267-276 https://doi.org/10.3354/meps10084
Fauchald, P., Anker-Nilssen, T., Barrett RT, Bustnes, J.O., Bårdsen, B.-J., Christensen-Dalsgaard, S., Descamps, S., Engen, S., Erikstad, K.E., Hanssen, S.A., Lorentsen, S.-H., Moe, B., Reiertsen, T.K., Strøm, H., Systad, G.H. (2015). The status and trends of seabirds breeding in Norway and Svalbard. NINA Report 1151: 84 pp.
Gaston, A.J. og Jones, I.L. 1998. The auks: Alcidae. Oxford University Press Inc. New York.
Merkel, B., & Strøm, H. (2024). Post‐colony swimming migration in the genus Uria. Journal of Avian Biology, 2024(1-2), e03153.
Mesquita MS, Erikstad KE, Sandvik H, Barrett RT, Reiertsen TK, Anker-Nilssen T, Hodges KI and Bader J (2015). There is more to climate than the North Atlantic Oscillation: a new perspective from climate dynamics to explain the variability in population growth rates of a long-lived seabird. Front. Ecol. Evol. 3:43. doi: 10.3389/fevo.2015.00043
Sandvik, H., Barrett, R.T., Erikstad, K.E., Myksvoll, M.S., Vikebø, F., Yoccoz, N.G., Anker-Nilssen, T., Lorentsen, S.H., Reiertsen, T.K., Skarðhamar, J. and Skern-Mauritzen, M., 2016. Modelled drift patterns of fish larvae link coastal morphology to seabird colony distribution. Nature communications, 7(1), p.11599.
Stokke BG, Dale S, Jacobsen K-O, Lislevand T, Solvang R og Strøm H (24.11.2021). Fugler: Vurdering av lomvi Uria aalge for Norge. Rødlista for arter 2021. Artsdatabanken. https://lister.artsdatabanken.no/rodlisteforarter/2021/28793. Nedlastet 09.03.2026.
Wassmann, P., M. Reigstad, T. Haug, B. Rudels, M.L. Carroll, H. Hop, G.W. Gabrielsen, S. Falk-Petersen, S.G. Denisenko, E. Arashkevich, D. Slagstad, and O. Pavlova. 2006. Food web and carbon flux in the Barents Sea. Progress on Oceanography 71: 232 – 287.